一文速覽丨2019年曹雪濤團隊在Science,PNAS發表成果盤點
2019年,曹雪濤團隊在Science,Nature Immunology,PNAS 等雜志上發表了13篇重要研究成果,在免疫學領域取得重大進展,iNature系統盤點一下曹雪濤團隊的研究成果:
【1】干擾素-γ(IFN-γ)對于細胞內細菌固有的免疫反應至關重要。 非編碼RNA和RNA結合蛋白(RBP)在研究巨噬細胞中IFN-γ激活的信號通路的調控中不是很清楚。2019年11月18日,曹雪濤團隊在Nature Immunology 在線發布題為“Inducible degradation of lncRNA Sros1 promotes IFN-γ-mediated activation of innate immune responses by stabilizing Stat1 mRNA”的研究論文,該研究發現microRNA miR-1通過降解胞質長非編碼RNA Sros1,間接穩定Stat1信使RNA來促進巨噬細胞中IFN-γ介導的李斯特菌的清除。 Sros1的可誘導降解或遺傳缺失導致先天免疫應答增強的IFN-γ依賴性激活。 這些發現揭示了復雜的RNA-RNA調控網絡,參與了宿主-病原體相互作用中細胞因子引發的先天反應。
【2】宿主細胞代謝可以通過病毒感染來調節,從而影響病毒存活或清除。由病毒 ?-宿主相互作用中的N6-甲基腺苷(m6A)修飾介導的細胞代謝重新布線仍然是未知的。2019年8月22日,曹雪濤團隊在Science在線發表題為”N6-methyladenosine RNA modification–mediated cellular metabolism rewiring inhibits viral replication“的研究論文,該研究發現,為了響應病毒感染,宿主細胞損害RNA m6A去甲基酶ALKBH5的酶活性。 這增加了α-酮戊二酸脫氫酶(OGDH)mRNA的m6A甲基化,從而降低了其mRNA穩定性和蛋白質表達。 總而言之,該研究顯示OGDH和衣康酸以先天免疫非依賴性方式促進病毒復制,提出靶向OGDH-衣康酸代謝反應以控制病毒感染性疾病。
【3】DNA病毒通常在進入后將基因組DNA排出到宿主細胞的細胞核中。然而,尚不清楚核病原體衍生的DNA如何觸發先天免疫反應。 2019年7月18日,曹雪濤團隊在Science 在線發表題為“Nuclear hnRNPA2B1 initiates and amplifies the innate immune response to DNA viruses”的研究論文,該研究報道異質核核糖核蛋白A2B1(hnRNPA2B1)識別致病性DNA并擴增IFN-α/β的產生。因此,hnRNPA2B1在啟動IFN-α/β產生和增強STING依賴性細胞質抗病毒信號傳導中起重要作用。
【4】2019年6月25日,曹雪濤團隊在PNAS 在線發表題為“An endosomal LAPF is required for macrophage endocytosis and elimination of bacteria”的研究論文,該研究發現LAPF(含有PH和FYVE結構域的溶酶體相關和凋亡誘導蛋白)是細胞膜內陷介導的內吞作用所必需的。Lapf缺陷型巨噬細胞具有內吞和消除細菌的能力受損。巨噬細胞特異性Lapf缺陷小鼠對大腸桿菌(E.coli)感染更敏感,細菌負荷更高。此外,Lapf缺乏損害TLR4內吞作用,導致TLR-觸發的促炎細胞因子的產生減弱。LAPF定位于早期內涵體并與caveolin-1相互作用。酪氨酸激酶Src對LAPF的磷酸化是LAPF-Src -Caveolin復合物形成和內吞作用以及細菌消除所必需的。總的來說,該研究表明LAPF對細菌的內吞作用和誘導炎癥反應至關重要,這表明LAPF和Src可能是控制傳染病的潛在靶標;
【5】2019年6月18日,曹雪濤團隊在Cell Research 在線發表題為“Interferon-inducible cytoplasmic lncLrrc55-AS promotes antiviral innate responses by strengthening IRF3 phosphorylation”的研究論文,該研究鑒定了一種新的細胞質長非編碼RNA(lncRNA),lncLrrc55-AS,它驅動正反饋環以促進干擾素調節因子3(IRF3)信號傳導和IFN-I產生;
【6】2019年5月10日,中國醫學科學院基礎醫學研究所北京協和醫學院基礎學院曹雪濤,姜明紅等人在Cellular & Molecular Immunology 在線發表題為“E3 ubiquitin ligase RNF170 inhibits innate immune responses by targeting and degrading TLR3 in murine cells”的研究論文,該研究通過免疫沉淀巨噬細胞中的TLR3結合蛋白,將環指蛋白170(RNF170)鑒定為結合TLR3的E3連接酶。RNF170介導K766在TLR3的TIR結構域中的K48連接的多泛素化,并通過蛋白酶體途徑促進TLR3的降解。RNF170的遺傳缺失在體外和體內選擇性地增強TLR3觸發的先天免疫應答。 研究結果揭示了RNF170通過促進TLR3降解選擇性抑制TLR3觸發的先天免疫反應的新作用;
【7】2019年4月29日,浙江大學醫學院曹雪濤團隊在Nature Immunology期刊上在線發表題為“The long noncoding RNA Lnczc3h7a promotes a TRIM25-mediated RIG-I antiviral innate immune response”的研究論文,該研究確定了一種新的長非編碼RNA(Lnczc3h7a),它與TRIM25結合并促進RIG-I介導的抗病毒先天免疫反應。 在機制上,Lnczc3h7a與TRIM25和活化的RIG-I結合,在病毒感染的早期階段用作穩定RIG-I-TRIM25復合物的分子支架。Lnczc3h7a促進TRIM25介導的RIG-1的K63連接的泛素蛋白化,從而促進下游信號轉導。研究結果表明,宿主RNA可以增強先天免疫傳感器對外來RNA的反應,確保有效的抗病毒防御;
【8】N6-甲基腺苷(m6A)修飾通過調節mRNA生物學在各種細胞反應中起重要作用。然而,m6A修飾如何通過影響免疫轉錄物的翻譯而與先天免疫有關還有待進一步研究。2019年4月23日,曹雪濤團隊在Nature Communications在線發表題為“Mettl3-mediated mRNA m6A methylation promotes dendritic cell activation”的研究論文,該研究發現RNA甲基轉移酶Mettl3介導的mRNA m6A甲基化促進樹突細胞(DC)激活和功能。從機制上講,Mettl3介導的CD40,CD80和Tirap的m6A增強了它們在DC中的翻譯,用于刺激T細胞活化,并增強TLR4 / NF-κB信號傳導誘導的細胞因子產生。該研究結果確定了Mettl3介導的m6A修飾在增加某些免疫轉錄物的翻譯中的新作用,用于生理促進DC活化和基于DC的T細胞應答;
【9】2019年3月27日,曹雪濤院士團隊在《自然免疫學綜述》(Nature Reviews Immunology)(IF41.98)上發表了長達16頁的題為“Epigenetic regulation of the innate immune response to infection” 的綜述文章;
【10】2019年3月6日,曹雪濤等人在Journal of Experimental Medicine雜志在線發表題為“KAT8 selectivelyinhibits antiviral immunity by acetylating IRF3”的文章,該文章揭示了KAT8和IRF3賴氨酸乙酰化在抑制抗病毒先天免疫中的關鍵作用;
【11】2019年2月26日出版的新一期《免疫學》(Immunity)雜志刊登了中國工程院院士、南開大學校長曹雪濤研究團隊題為“CCR7 Chemokine Receptor-Inducible lnc-Dpf3 Restrains Dendritic Cell Migration byInhibiting HIF1a-Mediated Glycolysis”的研究論文。該研究報道了一種稱為lnc-Dpf3的新型長鏈非編碼RNA能通過抑制樹突狀細胞(DC)體內遷移進而抑制炎癥性疾病的發生發展。這也是2019年曹雪濤團隊繼Nature Medicine、Cellular & Molecular Immunology的第三篇重要研究成果;
【12】2019年1月15日,曹雪濤團隊在Nature Medicine在線發表題為“Tumor-educated B cells selectively promote breast cancer lymph node metastasis by HSPA4-targeting IgG”的研究論文,該論文使用乳腺癌自發淋巴結轉移的小鼠模型,顯示原發性腫瘤在draining淋巴結中誘導B細胞積聚。這些B細胞通過產生靶向糖基化膜蛋白HSPA4的致病性IgG,選擇性地促進淋巴結轉移,并且在腫瘤細胞中激活HSPA4結合蛋白ITGB5和下游Src / NF-κB途徑用于CXCR4 /SDF1α軸介導的轉移。研究結果確定了B細胞及其衍生抗體在淋巴結轉移前生態位形成中的關鍵作用,為癌癥干預提供了潛在的靶點;
【13】2019年1月11日,曹雪濤團隊在Cellular & Molecular Immunology雜志在線發表題為“Intracellular HSP70L1 inhibits human dendritic cell maturation by promoting suppressive H3K27me3 and H2AK119Ub1 histone modifications”的研究論文,該論文證明細胞內HSP70L1通過抑制MHC和共刺激分子表達來抑制人DC成熟,與細胞外HSP70L1的佐劑活性相反。細胞內HSP70L1的穩定性依賴于已知的表觀遺傳調節因子DNAJC2。該研究結果為細胞內HSP70L1對細胞發育的表觀遺傳調控提供了新的見解;
1

先天性免疫反應可保護宿主免受病原體感染,并向病原體施加進化壓力,以減弱這些反應并確保其存活和復制的策略。這些不斷變化的壓力導致了跨宿主-病原體相互作用的先天性免疫穩態的復雜機制,但尚未得到全面了解。尤其是,更好地了解控制宿主與病原體相互作用并促進入侵病原體清除或逃逸的調節劑,可以確定傳染病和慢性炎癥的治療靶標。
非編碼RNA(ncRNA)調節各種先天和適應性免疫過程。 MicroRNA(miRNA)是研究最深入的ncRNA,通常在免疫反應中充當基因表達的轉錄后調節劑,而長的非編碼RNA(lncRNA)已成為各種生理學中基因表達的其他關鍵調節因子。 LncRNA通過多種機制調節免疫反應和炎癥中的信號傳導途徑,包括充當調節蛋白的向導,支架或誘餌。
核lncRNAs通過支架亞核結構和指導核小體定位來調節免疫反應基因的轉錄。LncRNA–ACOD1直接與其酶伴侶谷氨酸–草酰乙酸轉氨酶2結合,以增強該酶的催化活性,從而促進病毒感染。盡管有這些觀察結果,目前尚不清楚調控宿主與病原體相互作用的lncRNA的范圍和設置。此外,lncRNA用來調節針對細菌感染的先天免疫應答的多種機制使得難以預測是否以及如何涉及lncRNA。
單核細胞增生李斯特菌是一種細胞內致病性革蘭氏陽性細菌,是研究宿主與病原體相互作用和細胞內感染的理想模型。單核細胞增生李斯特氏菌被易感的真核宿主攝取后,便會轉變成病原體,以在惡劣的環境中生存。感染的命運取決于巨噬細胞活化與病原體抵抗宿主細胞殺菌能力之間的平衡。 II型干擾素(IFN-γ)通過信號轉換器JAK和轉錄激活因子STAT發出信號,從而具有有效的抗細胞內微生物活性,該轉錄因子可激活巨噬細胞以對抗細胞內微生物。
為了解決STAT1的數量如何在免疫反應中達到平衡,該研究篩選了單核細胞增生李斯特菌和巨噬細胞之間相互作用的miRNA。研究人員發現,miR-1促進了巨噬細胞中針對細胞內單核細胞增生李斯特菌感染的IFN-γ介導的先天應答。 miR-1在轉錄后增加Stat1 mRNA的表達,并增強巨噬細胞中的IFN-γ信號傳導。在無偏差的miRNA體內沉淀(miRIP)方法中,發現lncRNA Sros1(STAT1的抑制性非編碼RNA)被miR-1靶向。 miR-1介導的Sros1降解可穩定Stat1 mRNA,并促進IFN-γ-STAT1介導的先天應答。
總而言之,這些發現揭示了復雜的RNA-RNA調控網絡,參與了宿主-病原體相互作用中細胞因子引發的先天反應。
參考消息:
https://www.nature.com/articles/s41590-019-0542-7
2
細胞代謝涉及各種生物過程,包括病毒 ?-宿主相互作用。在識別或感知入侵病毒后,宿主細胞主動地重新編程代謝,以提供病毒存活的必要組分或抑制病毒復制以進行清除。多個監管機構在病毒 ?-宿主互動期間參與這些變化。病毒還可以調節宿主代謝途徑以完成其生命周期,逃避宿主免疫應答并為持續感染創造有利的細胞微環境。例如,腺病毒感染通過誘導Warburg樣轉變為有氧糖酵解來改變宿主細胞代謝,從而合成病毒復制所需的蛋白質和核酸。
ALKBH5在體外和體內促進病毒復制,而與先天反應無關
RNA修飾,尤其是最常見的哺乳動物mRNA修飾m6A,可以調節基因表達并調節病毒感染。例如,m6A甲基轉移酶復合物成分METTL3 / 14限制黃病毒Zika的產生,而m6A脫甲基酶ALKBH5和FTO增強其產量。
宿主細胞損害ALKBH5介導的RNA m6A去甲基化以抑制病毒復制
在該研究,研究人員報告,響應病毒感染,宿主細胞損害RNA m6A去甲基酶ALKBH5的酶活性。 這增加了α-酮戊二酸脫氫酶(OGDH)mRNA的m6A甲基化,從而降低了其mRNA穩定性和蛋白質表達。減少的OGDH減少了病毒復制所需的代謝物衣康酸的產生。隨著體內OGDH和衣康酸產生減少,ALKBH5缺陷小鼠顯示出對病毒攻擊的先天免疫應答依賴性抗性。該研究結果表明,m6A RNA修飾介導的OGDH-衣康酸途徑的下調重新編程細胞代謝以抑制病毒復制,提出了控制病毒感染的潛在靶標。
ALKBH5直接消除OGDH mRNA上的m6A修飾,以增加其穩定性和表達
總而言之,該研究顯示OGDH和衣康酸以先天免疫非依賴性方式促進病毒復制,提出靶向OGDH-衣康酸代謝反應以控制病毒感染性疾病。
參考信息:
https://science.sciencemag.org/content/early/2019/08/21/science.aax4468?rss=1
3
通過模式識別受體(PRR)識別病毒核酸可以觸發對病毒的宿主先天免疫應答。這導致由干擾素調節因子(IRF)信號傳導介導的由NF-κB信號傳導和I型干擾素調節的促炎細胞因子的產生。通常,一旦DNA病毒進入宿主細胞,它們就會在宿主細胞核內彈出并復制它們的基因組DNA。然而,在細胞核內識別病原體的DNA的過程仍然是個謎。
hnRNPA2B1激活抗病毒防御以抑制DNA病毒復制
迄今為止,只有一種蛋白質γ-干擾素誘導蛋白16(IFI16)被提出用于識別細胞核內的病毒DNA并激活IFN-I的產生和炎癥反應。鑒于宿主細胞經常遇到核病原體衍生的DNA,因此該研究試圖鑒定細胞核內其他未表征的IFN-1引發劑。
hnRNPA2B1的二聚化是其核質轉運和活化所必需的
已經鑒定出許多能夠識別病毒DNA并誘導IFN-α/β產生的蛋白質,如RNA聚合酶III,IFI16,干擾素調節因子(DAI)的DNA依賴性激活劑,LRRFIP1,LSm14A和環狀GMP-AMP(cGAMP)合酶(cGAS) )等。然而,只有細胞質cGAS和DNA-PK在功能上被證實為體內DNA傳感器。還報道了幾種蛋白質參與DNA病毒誘導的炎癥反應,包括黑素瘤2(AIM2),IFI16,Rad50和Sox2等。因此,需要更全面地了解針對DNA病毒的先天免疫應答,尤其是關于將病原體衍生的DNA的核識別與細胞質信號傳導的激活聯系起來的途徑。
hnRNPA2B1是cGAS,IFI16和STING有效的I型干擾素誘導所必需的
在該研究,檢查了與單純皰疹病毒-1(HSV-1)的基因組DNA結合的核蛋白,以及在病毒感染后易位至細胞質。該分析揭示了異質核核糖核蛋白A2B1(hnRNPA2B1)作為限制DNA病毒感染的I型干擾素產生的核啟動子。在直接識別核病原體衍生的DNA后,hnRNPA2B1易位至細胞質以啟動先天免疫應答。然后,hnRNPA2B1同時促進mRNA的核質轉運和細胞質表達,例如CGAS,IFI16和STING mRNA,其擴增抗病毒先天免疫信號。總之,這項研究強烈表明,核DNA傳感器如hnRNPA2B1是先天免疫防御的重要貢獻者。
參考信息:
https://science.sciencemag.org/content/early/2019/07/17/science.aav0758
4
先天免疫細胞檢測入侵病原體并發出適當的炎癥反應以消除感染。這些細胞表達多種模式識別受體(PRR),如Toll樣受體(TLRs),它們可識別病原體相關分子模式(PAMPs)并激活下游信號級聯以誘導炎癥。 TLR4是一種特異性識別革蘭氏陰性菌脂多糖(LPS)的PRR,通過骨髓分化因子88(MyD88)激活NF-κB和絲裂原活化蛋白激酶(MAPK),進而誘導促炎細胞因子的表達。此外,TLR4在識別LPS時被內化到內體中,通過銜接蛋白Toll-IL-1抗性結構域誘導IFN-β(TRIF)和TRAM(TRIF相關銜接子),誘導IRF3依賴性I型IFN產生。然而,先天信號傳導誘導有效的先天反應的分子機制需要進一步研究。
Lapf缺陷型巨噬細胞內吞較少的大腸桿菌和金黃色葡萄球菌,并消除較少內吞的大腸桿菌
先天免疫細胞,例如巨噬細胞,樹突細胞(DC),單核細胞和嗜中性粒細胞,可通過內吞作用內化入侵病原體。在內化后,病原體成為從早期內體到溶酶體的細胞器的一系列囊泡運輸的靶標,其中它們被Mst1-Mst2-Rac信號傳導誘導的活性氧物質(ROS)殺死并隨后被水解酶降解。然后將來自這些病原體的抗原呈現在主要組織相容性復合物(MHC)分子上,其隨后被T細胞受體識別并激活適應性免疫系統的CD4 +和CD8 + T細胞。內吞作用通過多種機制發生,特別是網格蛋白介導的內吞作用,細胞膜內陷介導的內吞作用,巨胞飲作用和吞噬作用。然而,仍然需要充分研究將入侵病原體的內吞作用與先天信號傳導的激活聯系起來的機制。
Lapf缺陷型小鼠對大腸桿菌感染更敏感,炎性細胞因子較少
Caveolae是一種專門的脂筏,是20世紀50年代首次描述的球形質膜內陷。從那以后,細胞膜內陷廣泛參與許多細胞過程,如內吞作用,轉胞吞作用(一種特殊形式的內吞作用),脂質穩態和信號轉導。細胞膜內陷也參與受體的內吞作用,如TLR4,這是先天免疫反應和信號轉導的必要調節機制。細胞膜內陷的主要成分蛋白質是caveolins和cavins。 Caveolin家族由三個成員組成,即caveolin-1(Cav1),caveolin-2(Cav2)和caveolin-3(Cav3)。 Cav1和Cav2在大多數細胞類型中表達。 Cav3在肌肉細胞中特異性表達。細胞膜內陷中的Cav1結合TLR4,內皮型一氧化氮合酶(eNOS),MAPK,環加氧酶(COX)和整合素信號分子,啟動不同的信號通路。然而,Cav1在細菌感染中的先天功能及其潛在機制尚未確定。
Src-LAPF-Cav1復合物形成的破壞損害了細菌的內吞作用
該研究發現LAPF(含有PH和FYVE結構域的溶酶體相關和凋亡誘導蛋白)是細胞膜內陷介導的內吞作用所必需的。 Lapf缺陷型巨噬細胞具有內吞和消除細菌的能力受損。巨噬細胞特異性Lapf缺陷小鼠對大腸桿菌(E.coli)感染更敏感,細菌負荷更高。此外,Lapf缺乏損害TLR4內吞作用,導致TLR-觸發的促炎細胞因子的產生減弱。LAPF定位于早期內涵體并與caveolin-1相互作用。酪氨酸激酶Src對LAPF的磷酸化是LAPF-Src -Caveolin復合物形成和內吞作用以及細菌消除所必需的。總的來說,該研究表明LAPF對細菌的內吞作用和誘導炎癥反應至關重要,這表明LAPF和Src可能是控制傳染病的潛在靶標。
參考信息:
https://www.pnas.org/content/116/26/12958.long
5
先天免疫反應是宿主防御入侵病毒的第一道防線。先天受體識別病毒成分并激活下游信號,包括干擾素調節因子3(IRF3)和核因子-κB(NF-κB)途徑。轉錄因子IRF3的激活誘導I型干擾素(IFN-1)的產生,其在宿主防御病毒入侵中起關鍵作用。上游信號分子(例如,TBK1,IKKε)磷酸化IRF3,引發其二聚化和易位,它可以刺激IFN-1的表達。蛋白磷酸酶2A(PP2A)使IRF3去磷酸化,從而負調節IFN-I的產生。如何精確平衡IRF3的磷酸化和去磷酸化以確保適當的IFN-I產生,同時避免組織先天免疫反應期間的損害尚不完全清楚。
已知表觀遺傳調節因子在免疫細胞功能的調節以及免疫疾病的發病機制中發揮重要作用。長的非編碼RNA(lncRNA)正在成為先天和適應性免疫的關鍵調節因子。 一些lncRNA直接與染色質修飾因子,異質核核糖核蛋白(hnRNPs)或轉錄因子相互作用以調節免疫相關基因的轉錄,而其他lncRNA形成多亞基復合物以調節先天免疫應答途徑。細胞質lncRNA已顯示通過調節翻譯后修飾(PTM)或細胞代謝來控制信號組分的活性。此外,一些差異表達的lncRNA調節炎癥先天反應和病原體逃避或宿主期間的存活 ?-病原體相互作用。 LncRNAs還可以與信號分子相互作用,以控制不同的生物過程。
在先前的研究中,病毒感染誘導IFN-I依賴性lncRNA lnc-lsm-3b和IFN-I非依賴性lncRNA lncRNA-ACOD1的表達,其通過不同機制調節先天反應和病毒復制。該研究鑒定了一種新的細胞質長非編碼RNA(lncRNA),lncLrrc55-AS,它驅動正反饋環以促進干擾素調節因子3(IRF3)信號傳導和IFN-I產生。
該研究顯示lncLrrc55-AS通過IFN-JAK-STAT途徑在多種細胞類型中被病毒誘導。LncLrrc55-AS缺陷型小鼠顯示出弱化的抗病毒免疫應答,并且更容易受到病毒攻擊。機制上,lncLrrc55-AS結合磷酸酶甲酯酶1(PME-1),并促進PME-1與磷酸酶PP2A(IRF3信號傳導的抑制劑)之間的相互作用。LncLrrc55-AS支持PME-1介導的去甲基化和PP2A的失活,從而增強IRF3磷酸化和信號傳導。PME-1表型缺失,發現lncLrrc55-AS缺失,導致IRF3磷酸化和IFN-I產生減少。總而言之,該研究鑒定出IFN誘導的lncRNA作為IFN-1產生的正調節因子,增加了對先天免疫和炎癥中lncRNA介導的信號傳導調節的機制洞察。
參考信息:
https://www.nature.com/articles/s41422-019-0193-0
6
作為Toll樣受體(TLR),TLR3識別雙鏈RNA(dsRNA)并在先天免疫反應中起關鍵作用。 TLR3由富含亮氨酸的重復序列(LRR),跨膜結構域和 toll/interleukin-1受體(TIR)結構域。活性TLR3位于內質網(ER)中并保持閉合構象,使其TIR結構域不能被細胞質蛋白質所接近。在識別dsRNA后,TLR3經歷從ER到內溶酶體的易位;然后,dsRNA與內溶酶體中的LRR結構域結合,引起TLR3的構象變化和二聚化;緊接著,細胞質中的TIR結構域募集銜接蛋白TRIF,啟動下游IRF3和NF-κB信號傳導,從而導致I型干擾素(IFN)和炎性細胞因子的轉錄。
RNF170直接與TLR3相互作用
已發現TLR3在控制病毒感染方面具有保護作用,另外TLR3缺乏與嚴重的傳染病,如皰疹病毒性腦炎有關。然而,TLR3的過度表達也可能由于TLR3介導的炎癥反應而引起一些自身免疫性疾病。因此,精確調節TLR3的表達功能對維持免疫穩態和預防相關疾病至關重要。但是,調節TLR3表達和功能的詳細機制尚不清楚。
泛素化是在翻譯后水平改變蛋白質表達或功能的重要方法,并且已發現其在許多階段參與先天免疫應答的調節。附著于底物的泛素化蛋白也可以形成不同的多鏈接并通過不同的機制影響底物的表達或功能。 K48連接的多聚泛素化通過蛋白酶體途徑介導底物的降解,而K63連接的多泛素化促進底物的活化。據報道,許多E3連接酶參與TLR3下游信號分子的調節。但是泛素化對TLR3表達或功能以及相關的TLR3結合的E3連接酶的潛在調節作用仍然很大程度上未知。
在本研究中,通過免疫沉淀巨噬細胞中的TLR3相互作用蛋白,研究人員將環指蛋白170(RNF170)鑒定為結合TLR3的E3連接酶。RNF170之前被鑒定為位于ER膜的E3連接酶,其介導-三磷酸肌醇受體的泛素化和降解。然而,尚未鑒定RNF170在先天免疫應答中的底物和功能。在這里,研究人員證明了RNF170促進了TLR3中K766的K48連接多聚泛素化,這導致TLR3通過蛋白酶體途徑降解。因此,RNF170抑制TLR3信號傳導并抑制TLR3觸發的先天免疫應答。
參考信息:
https://www.nature.com/articles/s41423-019-0236-y
7
RNA傳感器RIG-I(視黃酸誘導基因I)在宿主防御反應中起著至關重要的作用,可以消除入侵的RNA病毒。感染后,RIG-I C末端結構域識別并結合攜帶5'-三磷酸(5'-ppp RNA)或5'-二磷酸(5'-pp RNA)的病毒RNA,然后激活至啟動下游先天信號傳導以誘導I型干擾素(IFN)。RIG-I的不同泛素修飾調節抗病毒免疫反應,例如,E3泛素蛋白連接酶c-Cbl和CHIP通過K48連接的泛素蛋白化促進RIG-I的蛋白酶體降解,其抑制RIG-I依賴性抗病毒信號傳導。然而,E3連接酶TRIM25在其兩個N-末端半胱天冬酶激活和募集結構域(CARD)中介導RIG-1的K63連接泛素化。K63-泛素蛋白化的RIG-I形成與線粒體抗病毒信號蛋白(MAVS)相互作用的寡聚體,以引發下游信號級聯并引發針對RNA病毒的先天免疫應答,包括I型IFNs的產生。
盡管TRIM25在RIG-I介導的抗病毒反應中起關鍵作用,但調節其抗病毒活性的機制卻知之甚少。研究表明,TRIM25受RNA結合調節。登革熱亞基因組RNA與TRIM25結合并阻止TRIM25去泛素化,從而負調節RIG-I抗病毒信號通路,使病毒能夠逃避宿主抗病毒先天免疫反應。然而,宿主RNA是否通過TRIM25影響抗病毒先天免疫仍不清楚。
長非編碼RNA(lncRNA)通過不同的機制在免疫細胞發育和免疫反應中發揮重要作用,包括信號,誘餌,指導或支架作用。先前已經發現了通過直接結合靶蛋白而參與免疫細胞發育和病毒感染的幾種lncRNA,例如,lnc-DC通過直接結合促進STAT3磷酸化,因此是樹突細胞分化所必需的。然而,在先天免疫應答的早期階段與關鍵先天信號分子相互作用以增強抗病毒先天免疫應答的lncRNA需要進一步鑒定。
在這里,研究人員鑒定了先前未描述的lncRNA:Lnczc3h7a,其與TRIM25和活化的RIG-I結合以穩定TRIM25-RIG-I相互作用。有趣的是,Lnczc3h7a在抗病毒反應的早期階段作為這種相互作用的分子支架,增強了RIG-I和下游信號傳導的K63連接泛素化。結果,Lnczc3h7a增加I型IFN產生并支持針對RNA病毒感染的更有效的先天免疫應答。這些發現揭示了宿主(自身)lncRNA在進入外源(非自身)RNA和抗病毒先天免疫應答的強度后,促進先天傳感器活化的先前未被認可的作用。
參考信息:
https://www.nature.com/articles/s41590-019-0379-0
8
mRNA的轉錄后修飾,包括N6-甲基腺苷(m6A),5-甲基胞嘧啶(m5C)和假尿苷(ψ),參與多個細胞過程的表觀遺傳調控,在影響mRNA穩定性,翻譯等過程。作為對哺乳動物mRNA最普遍的轉錄后修飾,m6A的系統和動態調節由多個writer/eraser組分協調。它主要由含有甲基轉移酶樣(METTL)酶METTL3和METTL14,Wilms腫瘤相關蛋白(WTAP)的大RNA甲基轉移酶復合物催化,其中METTL3負責催化m6A形成; 并通過兩種去甲基化酶去除:FTO和α-酮戊二酸依賴性雙加氧酶AlkB同系物5(ALKBH5)。
m6A修飾水平在DC成熟中增加
最近,確定了幾個屬于METTL3-METTL14-WTAP復合體的新成員;據報道RBM15 / RBM15B將該復合物募集到某些mRNA和lncRNA XIST以促進m6A形成;鑒定KIAAI1429和ZC3H3調節m6A甲基化并在不同細胞中發揮不同的功能。此外,一組m6A readers,如YTH-結構域家族(YTHDF)1和YTHDF2,可識別m6A,并分別促進m6A修飾的mRNA的翻譯和降解。新出現的證據表明,m6A修飾通過介導RNA衰變來調節多種生物途徑,如干細胞分化,腫瘤發生和病毒復制。最近,m6A被證明是宿主免疫細胞區分自身和非自身的重要機制,也可能被病毒劫持以逃避免疫反應;然而,m6A在先天免疫中的這種作用不能用RNA降解機制來解釋。因此,需要進一步研究m6A是否通過RNA降解獨立機制在哺乳動物細胞中,特別是在免疫細胞中具有其他生理功能。
Mettl3以m6A催化活性依賴性方式促進DC成熟
樹突狀細胞(DC)是連接先天性和適應性免疫應答的特異性抗原呈遞細胞(APC)。DC對于啟動適應性免疫應答以消除入侵病原體以及誘導對無害成分的免疫耐受以維持免疫穩態至關重要。DC對免疫激活或耐受的誘導嚴格依賴于不同成熟階段的不同DCs亞群。通常,未成熟DC(imDC)誘導免疫耐受,成熟DC(maDC)刺激并激活免疫應答,而調節DC(DCreg)通過抑制T細胞應答下調免疫應答。
Mettl3以m6A催化活性依賴性方式促進T細胞中的DC功能
眾所周知,不同階段DC激活的失調與多種炎癥,自身免疫和其他疾病密切相關。雖然已經深入研究了控制DC發育和功能的轉錄網絡,但表觀遺傳機制,特別是mRNA m6A修飾在該過程中的作用仍有待完全理解。因此,鑒定m6A甲基化在控制DC活化中的作用對于更好地理解免疫應答是至關重要的,并且還將具有重要的臨床意義。
Mettl3在DC成熟過程中增強先天反應和NF-κB信號傳導
可以通過LPS刺激上調的膜共刺激分子(包括CD40,CD80和CD86)直接調節DC在其抗原呈遞和T細胞活化中的功能和階段是公認的。然而,由于多個細胞可以對LPS刺激作出反應,因此仍然不清楚為什么只有專業的APC(例如maDC)在激活后高度且連續地表達這些共刺激分子以響應先天刺激。imDC,maDC和DCreg的分子譜分析是連續但不同的成熟和分化階段的三種DC,用于動態研究DC在不同亞群和階段的免疫激活和調節機制。
Mettl3促進體內Tirap,CD80和CD40 mRNA的翻譯
在本研究中,研究人員利用DC成熟和分化模型,包括骨髓來源的imDC,LPS刺激的BMDC(maDC)和DCreg,來研究m6A修飾在DC的成熟和功能中的表達模式和生物學作用。該研究證明mettl3在DC成熟過程中催化CD40,CD80及Tirap的m6A,并且提高它們的翻譯效率以促進DC活化和促進T細胞活化的功能。該研究揭示了通過m6A介導的相關免疫轉錄物甲基化對先天免疫的表觀遺傳調控。此外,該研究提供了一種新的機制,為什么APC一旦成熟和激活,優先表達更高水平的共刺激分子,以有效地啟動免疫反應。
參考信息:
https://www.nature.com/articles/s41467-019-09903-6
-
焦點事件
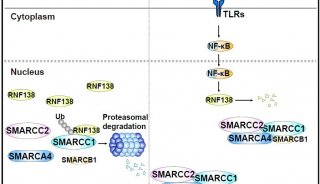
-
焦點事件

-
項目成果

-
焦點事件
-
焦點事件
-
科技前沿

-
焦點事件

-
焦點事件

-
焦點事件
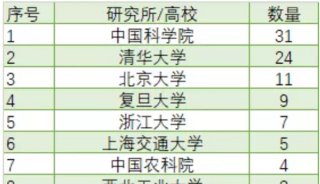
-
焦點事件

-
焦點事件

-
項目成果

-
焦點事件